Department of Sports Science
Research area VI - Motor Control and Training

Current projects
- Motor hysteresis as a proxy for motor planning

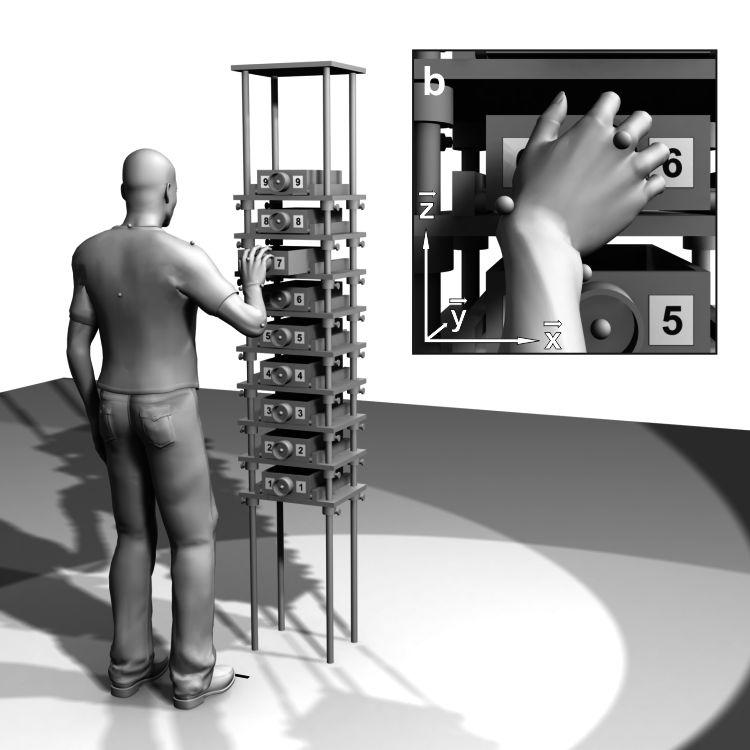
© Christoph Schütz - Universität Bielefeld In sequential reaching tasks, the grasp postures we select depend on our movement history (Rosenbaum & Jorgensen 1992). For example, when we open a column of slotted drawers in descending order, we use an overhand grasp for the highest drawer and persist on this grasp type for most of the subsequent drawers. In ascending sequences, we persist on our initially selected underhand grasp (Weigelt et al. 2009). This tendency to switch the movement state at different positions, depending on our movement history, is termed motor hysteresis (Kelso et al. 1994).
Experimental evidence suggests that motor hysteresis represents a cognitive aspect of motor planning (van der Wel et al. 2007). All motor planning results in a cognitive cost (Rosenbaum et al. 2007). In a sequential task, this cost is reduced by the reuse of a former motor plan. If posture selection is continuous (e.g., round drawer handles), each motor plan is created by a modification of the former plan, with a varying fraction of reuse (Schütz & Schack 2013a). A larger fraction of reuse reduces the cognitive cost, but increases the mechanical cost of the movement. Vice versa, an increase in mechanical cost reduces the fraction of reuse (Schütz & Schack 2016a).
The size of the motor hysteresis effect therefore not only serves as a proxy for the fraction of novel planning and plan reuse (larger fraction of novel planning = smaller hysteresis effect), it can also be used to estimate the cognitive and mechanical costs of a motor task. In the biomechanics lab, we use motor hysteresis as a tool to investigate the effect of different experimental perturbations on motor planning. The fraction of novel planning for arbitrary tasks can be estimated by a mathematical model of hysteresis (Schütz & Schack 2019a).- Motor primitives in reaching movements
© Christoph Schütz - Universität Bielefeld Many of the seemingly simple tasks we conduct in our daily lives, such as reaching for an object, require a series of sensorimotor transformations which map the object's position on our retina to an appropriate muscle activation pattern. A number of these transformations have infinitely many valid solutions: Each target position can be reached by various trajectories and all positions on a trajectory can be achieved by different postures. The selection of a single solution thus results in an ill-posed problem for the motor system (Jordan & Wolpert 1999).
One way to reduce redundancy on a postural level is to combine multiple independent joint angles into a motor primitive (Bernstein 1967). Joint angles in a motor primitive are no longer controlled individually but instead are coupled in their action. Previous studies on motor primitives for reaching movements were restricted to sagittal (Berret et al. 2009, Latash et al. 1995) or horizontal (Debicki & Gribble 2005, Sabatini 2002) target planes. However, it is unlikely that we have acquired specialized synergies for each arbitrary target plane. Instead, generalized synergies should exist to reach target locations within the three-dimensional workspace of the arm.
In the biomechanics lab, we investigate motor primitives of reaching movements in an augmented reality environment, with target locations distributed uniformly in the three-dimensional workspace of the arm (Schütz & Schack, 2013b). We also measure motor primitives in sequential reaching tasks (opening a column of drawers) and the motor hysteresis effect associated with each motor primitive. That way, we can test whether joint angles (1) have to be strictly controlled to satisfy the constraints of the task, or (2) can be controlled more loosely, to save cognitive cost (Schütz et al. 2016b).- Mutual interference of motor and working memory tasks
© Christoph Schütz - Universität Bielefeld Due to its cognitive demands, motor planning disrupts working memory in the same manner as other intervening cognitive tasks (Lawrence et al. 2001). This holds true even for the simplest reaching tasks (Logan & Fischman 2011). In a sequential reaching task, the disruptive effect was first demonstrated in our labs (Weigelt et al. 2009). Participants had to open a column of slotted drawers in a sequential order, retrieve a cup from each drawer, and memorize the letter inscribed on the bottom of the cup. Analysis of the verbal memory performance showed that the recency effect was eliminated by the concurrent motor task, which indicates a disruption of short term memory (Glanzer & Cunitz 1966).
As verbal and motor tasks share limited working memory resources (Cowan 2001), this disruptive effect should be mutual: Execution of a concurrent memory task should reduce the available cognitive resources for motor planning. In a number of experiments, we tested if verbal and spatial memory tasks would interfere with motor planning. The size of the motor hysteresis effect was used as a proxy for the fractions of motor planning and motor plan reuse (Schütz & Schack 2016a). All studies showed a disruptive effect of motor planning on verbal and spatial recall performance, but not vice versa. These findings indicate that motor planning has priority access to working memory resources and that interference effects are not mutual.An interesting result of our studies was that execution of a concurrent motor task only disrupted recency effects in a verbal but not in a spatial memory task (Schütz & Schack 2020b). Recency effects are commonly attributed to the episodic buffer (Baddeley et al., 1975), a non-domain specific memory subsystem of the central executive. The fact that recency effects in the verbal and spatial domain are affected differently by motor planning raises doubt on these established assumptions (Schütz & Schack 2020b). In the current project, we therefore test whether interference effects are only an effect of simultaneous encoding of two input streams from different domains or if the episodic buffer can no longer be considered non-domain specific.
- Subliminal social priming in a sports scenario
© Christoph Schütz - Universität Bielefeld The unconscious influence of external stimuli on human behavior has long been of interest and is still being debated (Molden 2014). The most common method to investigate a subliminal influence of stimuli on behavior is masked priming (Ansorge et al. 2014). In this paradigm, a masking stimulus follows a prime stimulus after a short delay. This backward masking (i.e., the mask following the prime) can prevent all conscious awareness of the prime. In this project, we ask whether a task-irrelevant but socially relevant cue, human gaze direction, can affect complex, whole-body responses automatically, that is, without conscious awareness, intentionality, and controllability.
Previous studies demonstrated that supraliminal gaze cues are processed even if detrimental to the task (Driver 1999), indicating that gaze processing is initiated without intention and cannot be suppressed by top-down strategic decisions. In contrast, subliminal gaze cues were only processed when presented in the context of supraliminal, informative primes or targets which resembled the supraliminal cues (Al-Janabi & Finkbeiner 2014). This supports the idea that supraliminal gaze cues were used for the intentional, top-down creation of action-trigger conditions (Kunde et al. 2003), which were then applied to the subliminal gaze cues unconsciously. Thus, subliminal gaze processing was not fully automatic but required top-down intentionality.
In the current project, we test participants in a controlled lab environment with high ecological validity (task resembling a sports scenario). Photorealistic cues (pictures of basketball players passing a ball past the left/right side of the participants) are combined with natural, full-body responses (blocking responses and sway on a force plate). First findings show that the processing of subliminal gaze cues is initiated without any intention, but can still be moderated (i.e., enhanced or suppressed) by top-down strategic decisions (Schütz et al. 2020c). Postural sway data further shows that subliminal primes actually initiate overt, full-body responses, whereas previous studies could only demonstrate differences in response times or in cortical activation (Dehaene et al. 1998).






Completed projects


- Cognitive mechanisms of motor planning - how cognitive and mechanical costs of a motor task affect the fractions of motor plan reuse and novel planning (DFG projectnumber 322322134)
Even the simplest voluntary movements require the creation of a motor plan. In sequential reaching tasks, motor plans are not created from scratch for every movement, but the former plan is partially reused. This reuse manifests in a persistence on the former posture, termed motor hysteresis. When a column of drawers is opened in sequential order, for example, people persist on a more pronated posture in the descending and on a more supinated posture in the ascending sequence. The motor hysteresis effect is explained by the cost-optimization hypothesis, which states that motor plans in a sequential task are created by a modification of the former plan, but with a varying fraction of reuse. A larger fraction of reuse reduces the cognitive cost of motor planning. At the same time, the larger persistence on the former, less optimal posture increases the mechanical cost of motor execution. The summed cost of the movement is minimized at an intermediate, optimal fraction of reuse.
The optimal fraction of reuse can be determined by a mathematical model. It depends on the relative weights assigned to the cognitive and mechanical cost of the task. Based on the mathematical model, one can predict how changes in the cognitive or mechanical cost of the task affect the fraction of reuse. An increase of the mechanical cost should decrease the optimal fraction of reuse and, in turn, the size of the hysteresis effect. This prediction has already been confirmed in a previous study of ours. In the proposed research program, we want to test whether the cost-optimization hypothesis also holds true for changes in the cognitive cost of a task.
To this end, we conduct two lines of experiments. In the first line, we experimentally increase the cognitive cost by depleting the available cognitive resources with a concurrent memory task. In the second line, we exploit intrinsic differences in the cognitive or mechanical cost, which are a result of hemispheric lateralization. Studies suggest that (1) the left hemisphere is dominant in motor planning and (2) the dominant hemisphere has superior control of limb dynamics. We therefore expect less cognitive cost of motor planning in the right limb and less mechanical cost of motor execution in the dominant limb, which should be reflected by according differences in hysteresis effect size between limbs.
If the cost-optimization hypothesis holds true, the underlying mathematical model can be used to estimate the relative weights assigned to the cognitive and mechanical cost during motor planning. To this end, we vary the mechanical cost of a two-parts movement sequence and measure the corresponding fractions of reuse. Based on this function, the weights of the two cost factors are determined by a model fit. This third line of experiments ultimately provides an estimate of how, during motor planning, 1 Nm of mechanical work is rated against the cognitive cost for the creation of a grasping movement plan from scratch.